 Экологические основы интегрированной защиты растений: учебник
Экологические основы интегрированной защиты растений: учебник
 ЭКОЛОГИЧЕСКИЕ ОСНОВЫ ИНТЕГРИРОВАННОЙ ЗАЩИТЫ РАСТЕНИЙ
ЭКОЛОГИЧЕСКИЕ ОСНОВЫ ИНТЕГРИРОВАННОЙ ЗАЩИТЫ РАСТЕНИЙ

 ГЛАВА 1. ПОНЯТИЕ О СИСТЕМАХ ИНТЕГРИРОВАННОЙ ЗАЩИТЫ РАСТЕНИЙ
ГЛАВА 1. ПОНЯТИЕ О СИСТЕМАХ ИНТЕГРИРОВАННОЙ ЗАЩИТЫ РАСТЕНИЙ


3.3.2. Межвидовые взаимодействия в биоценозах. Теория Триотрофа
Реализация экологических ниш и межвидовые отношения популяций экосистемы строго не детерминированы: виды могут вступать в новые отношения или менять их характер при изменении условий окружающей среды, перераспределяя затраты вещества и энергии на тактики жизнедеятельности.
Перераспределение в сообществах вещества и энергии между организмами позволяет им адаптироваться к изменившимся условиям и поддерживать сообщество в состоянии подвижного равновесия непрерывной сменой связей. Вредные организмы, будучи тесно связанными с растениями-хозяевами, входят в состав относительно постоянного флористического и фаунистического комплексов, формирующихся вокруг растений-хозяев как эдификаторов. В результате смены растений-эдификаторов усиливается гетерогенность сообществ вредных организмов разных агроэкосистем. Тем самым зеленые растения в структуре экосистем обеспечивают существование множества экологических ниш: возбудителей болезней и их антагонистов, фитофагов и энтомофагов, нейтральных и полезных для растений видов: эпифитов, микоризообразователей, симбионтов и др. Экологические ниши включают разнородные по видовому составу группы организмов. Так, экологическую нишу короедов на древесных растениях занимают и нематоды, экологическую нишу фитофагов сосущих соки растений – клопы, тли, вирусы. Экологическая ниша «стеблееда», «стволоеда» охватывает большую группу животных, среди которых многочисленны древоточцы, короеды, усачи. На листьях растений обитают эпифиты, возбудители болезней, фитофаги, а на корнях – микоризообразователи, ризосферные организмы, возбудители корневых гнилей, фитофаги (например, корневая свекловичная тля), фитонематоды.
По степени использования ресурсов среды различают "насыщенные" и "ненасыщенные" сообщества со свободными экологическими нишами в экосистемах. Более насыщенные сообщества формируются в почвенной среде, а ненасыщенные – в воздушно-наземной, с меньшей гетерогенностью популяций. |
Гетерогенность сообществ меняется в течение сезона и даже суток, так как в них включаются или исключаются новые виды вследствие их разной миграционной способности, суточного ритма жизненного цикла, гибели старых и воспроизводства новых особей в популяциях, а также открытости как агро-, так и естественных экосистем, исключая защищенный грунт. Гетерогенность сообществ определяется в значительной мере наличием или отсутствием в сообществе свободных экологических ниш, также общей емкостью среды.
В насыщенной почвенной среде свободные экологические ниши встречаются редко, а общая емкость среды меньше, чем воздушно-наземной, где существует экологический вакуум со свободными экологическими нишами.
Из ландшафтных сообществ состоят более крупные – зональные, географические. Их связь осуществляется пищевыми и пространственными отношениями популяций. Для ландшафтных сообществ характерна определенная структура, устойчивость, поддерживаемая взаимной регуляцией численности и отношений видов. Биогенный круговорот веществ объединяет все сообщества в единую биосферу Земли. Биоценозы рассматриваются как «сверхорганизмы», имеющие закономерные смены этапов – рождение, развитие, смерть – как любого живого существа, способного к внутренней регуляции системы.
Все многообразие популяций в сообществе можно разделить на три группы экологических эквивалентов по их функциональной роли в биологическом круговороте веществ в экосистемах. Эти группы занимают разные трофические уровни и вступают в разные трофические отношения друг с другом (рис. 10).

Рис. 10. Система триотрофа и потоки веществ и энергии через пищевые цепи экосистемы [30]
Первую группу составляют автотрофы, или продуценты (зеленые растения). Они создают органическое вещество из неорганических веществ в процессе фотосинтеза. Во вторую группу входят гетеротрофы (консументы), потребляющие эти вещества. Третью группу образуют редуценты (деструкторы), преобразующие и разлагающие органические вещества на более простые. Их биофильные элементы повторно используются растениями и вводятся ими в биогеохимические циклы (СО2, NH4, СО(NН2)2 и др.).
Существует примерно 500 тыс. видов автотрофов, занимающих 60 % суши, и примерно в 10 раз больше полностью зависящих от них видов гетеротрофов (консументов, редуцентов). Гетеротрофы получают необходимые им вещества от растений, а кислород, выделяемый растениями-автотрофами, делает возможным само существование жизни на суше.
Поддержание трофических связей в сообществах организмов и круговорот веществ и экосистемах зависят от постоянного притока солнечной энергии, образующейся в результате термоядерных реакций, идущих в недрах солнца. Количество энергии, посылаемой солнцем на землю, равно 13х1223 калорий в год. Каждый день солнце посылает такое количество энергии, которое эквивалентно примерно миллиону атомных бомб, аналогичных сброшенной на Хиросиму.
Односторонний приток солнечной энергии происходит в результате действия законов термодинамики. Первый закон термодинамики гласит, что энергия может превращаться из одной формы (например, света) в другую (например, потенциальную энергию химических связей органических соединений), но не может быть создана или уничтожена. Второй закон термодинамики утверждает, что не может быть ни одного процесса, связанного с превращением энергии, без потерь некоторой ее части. Эта часть превращается в недоступную тепловую энергию и рассеивается.
Треть солнечной энергии отражается Землей обратно в космическое пространство, а две трети поглощаются нашей планетой и превращаются в тепло. Лишь небольшая часть инсоляции (до 6 %) используется автотрофами для связывания углерода в процессе фотосинтеза.
Связывая лучистую энергию солнца в процессе фотосинтеза, растения накапливают ее в углеводах, представляющих собой потенциальную химическую энергию. Эта энергия поступает в биологический круговорот питательных веществ от растений (автотрофов) к гетеротрофам разного уровня, в том числе возбудителям инфекционных болезней растений и фитофагам. Вещества, циркулирующие во всех живых организмах, являются носителями энергии.
|
Перенос энергии и круговорот элементов включает сложную последовательность событий, в которой каждой трофической группе организмов (автотрофы, консументы, деструкторы) принадлежит строго определенная роль. При изменении хотя бы одной части нарушается равновесие, от которого зависит стабильность трофических связей и экосистемы в целом. |

Все вредные организмы включаются в биологический круговорот веществ – в цепи трофических связей. Например, колорадский жук входит в следующую трофическую цепь: картофель – колорадский жук – хищные клопы – певчие птицы – хищные птицы. Уничтожение популяции колорадского жука в агроэкосистеме нарушает трофические связи, обусловливая либо элиминацию связанных с вредителем видов, либо включение их в другие трофические цепи.
Для цепей питания характерны три свойства:
 каждая цепь состоит из небольшого (не более 5–6) числа звеньев. Это объясняется тем, что превращение веществ и энергии в экосистеме подчиняется «закону 10». Согласно этому закону только часть энергии и веществ (около 10 %), синтезируемых автотрофами, поступает на следующий трофический уровень, а из него – на следующий. Основная часть веществ и энергии (90 %) расходуется на поддержание жизнеспособности организмов соответствующего уровня трофической цепи. Тем самым передача веществ и энергии с одного трофического уровня на другой по схеме автотрофы – консументы – редуценты (деструкторы) осуществляется с низким коэффициентом полезного действия (КПД), что и объясняет ограниченное количество звеньев в трофической цепи, независимо от состава экосистемы;
каждая цепь состоит из небольшого (не более 5–6) числа звеньев. Это объясняется тем, что превращение веществ и энергии в экосистеме подчиняется «закону 10». Согласно этому закону только часть энергии и веществ (около 10 %), синтезируемых автотрофами, поступает на следующий трофический уровень, а из него – на следующий. Основная часть веществ и энергии (90 %) расходуется на поддержание жизнеспособности организмов соответствующего уровня трофической цепи. Тем самым передача веществ и энергии с одного трофического уровня на другой по схеме автотрофы – консументы – редуценты (деструкторы) осуществляется с низким коэффициентом полезного действия (КПД), что и объясняет ограниченное количество звеньев в трофической цепи, независимо от состава экосистемы;
 каждый последующий трофический уровень подчиняется правилу «величины особей и пирамиды чисел», которые различаются у хищников и паразитов. В трофической цепи хищников растет размер особей в популяции, но снижается их число, а в трофической цепи паразитов, наоборот, возрастает численность популяций и резко снижается размер особей. Например, численность злаковых тлей нередко достигает нескольких десятков тысяч на 1 м2. Более крупные тлевые коровки, поедающие тлей, встречаются в численности 5–7 особей на 1 м2, а насекомоядные птицы – лишь единично на 1 га. Ответвления от этой цепи составляют хищники и паразиты тлей, например златоглазки, которые тоже истребляются птицами или другими насекомыми. Для паразитов каждое последующее звено особей численно возрастает, а размеры особей уменьшаются. Так, личинки азиатской саранчи имеют размер тела 40 мм. Они заражаются личинками мух размером 9,5 мм, а личинки мух – наездниками размером около 5 мм;
каждый последующий трофический уровень подчиняется правилу «величины особей и пирамиды чисел», которые различаются у хищников и паразитов. В трофической цепи хищников растет размер особей в популяции, но снижается их число, а в трофической цепи паразитов, наоборот, возрастает численность популяций и резко снижается размер особей. Например, численность злаковых тлей нередко достигает нескольких десятков тысяч на 1 м2. Более крупные тлевые коровки, поедающие тлей, встречаются в численности 5–7 особей на 1 м2, а насекомоядные птицы – лишь единично на 1 га. Ответвления от этой цепи составляют хищники и паразиты тлей, например златоглазки, которые тоже истребляются птицами или другими насекомыми. Для паразитов каждое последующее звено особей численно возрастает, а размеры особей уменьшаются. Так, личинки азиатской саранчи имеют размер тела 40 мм. Они заражаются личинками мух размером 9,5 мм, а личинки мух – наездниками размером около 5 мм;
 цепи питания не замыкаются в себе, а всегда имеют ответвления. Ранее мы приводили пример с энтомофагами тли. Анализ цепей питания в экосистемах показывает, что биофаги (фитофаги, паразиты, хищники) активно влияют на этот процесс путем частичного разрушения предшествующего уровня, который служит для них источником энергии. Для последующих трофических уровней они создают специфический источник энергии путем преобразования поглощенных веществ: тканей собственного тела для биофагов, экскрементов – для копрофагов, трупов – для некротрофов, и тем самым пассивно определяют энергетический обмен у сменяющих их консументов. При этом используются не только сами организмы (их тела), но и продукты их метаболизма, выделяемые при дыхании, экскреции.
цепи питания не замыкаются в себе, а всегда имеют ответвления. Ранее мы приводили пример с энтомофагами тли. Анализ цепей питания в экосистемах показывает, что биофаги (фитофаги, паразиты, хищники) активно влияют на этот процесс путем частичного разрушения предшествующего уровня, который служит для них источником энергии. Для последующих трофических уровней они создают специфический источник энергии путем преобразования поглощенных веществ: тканей собственного тела для биофагов, экскрементов – для копрофагов, трупов – для некротрофов, и тем самым пассивно определяют энергетический обмен у сменяющих их консументов. При этом используются не только сами организмы (их тела), но и продукты их метаболизма, выделяемые при дыхании, экскреции.
Совместная и многосторонняя деятельность самых разных организмов экосистемы препятствует длительному накоплению мертвого вещества с заключенной в нем химической энергией.
Консументы, питающиеся остатками мертвых растений и животных, или мортмассой, (сапротрофы в самом широком значении этого термина), замедляют расход энергии тем, что используют часть потенциальной энергии, которая быстрее освобождалась бы без них, и тем самым жизнь в экосистемах становится более многообразной. Нематоды и членистоногие играют известную роль в регулировании роста грибов и бактерий, питаясь микроорганизмами и ограничивая их антибиотическое действие, которое могло бы подавлять рост и размножение микрофлоры и тем самым ускоряют поток энергии.
Существующий в экосистемах круговорот веществ создает возможность полной (в естественных экосистемах) или частичной (в агроэкосистемах) саморегуляции, придающей экосистемам относительную устойчивость. Состояние устойчивости обусловлено всеобщей трофической зависимостью (связью) организмов в сообществах, так как трофически независимых организмов не существует.
В круговороте энергии и веществ в любых типах экосистем действует принцип получения ресурсов и избавления от отходов в рамках круговорота элементов. Вследствие указанных процессов, присущих в той или иной степени всем экосистемам, вредные для автотрофов организмы вступают в определенные трофические связи друг с другом.
Помимо пищевых связей и параллельно им, в сообществах возникают пространственные отношения сожительствующих организмов. Особи одних видов используют других как среду обитания (эндопаразиты), как субстрат и место прикрепления (эктопаразиты и свободно живущие малоподвижные виды).
На базе пищевых и пространственных связей между видами возникают три основных типа отношений:
|



Возникновение указанных взаимоотношений зависит в значительной мере от характера экологических ниш, занимаемых видовыми популяциями.
Возможны три варианта взаимного пространственного расположения экологических ниш видовых популяций в сообществах экосистем:
 разделение (полное несовпадение);
разделение (полное несовпадение);
 частичное пересечение (перекрывание);
частичное пересечение (перекрывание);
 полное включение одной ниши в другую (рис. 11).
полное включение одной ниши в другую (рис. 11).

Рис. 11. Схематическое представление концепции экологической ниши [25]
Разделение ниш в экосистемах свидетельствует о существовании видов, приспособленных к различным экологическим условиям и не вступающих в конкуренцию друг с другом. В этом случае между популяциями складываются взаимоотношения типа нейтрализма, при которых они не связаны друг с другом пищевыми и пространственными отношениями. Примером является дивергенция, или расхождение экологических ниш, характерно для возбудителей почвенных и листо-стеблевых инфекций. Между ними отмечаются взаимоотношения нейтрализма, а также синергизма (греч synergeia – содружественное действие) при освоении экологических ниш. Так, возбудители септориоза интенсивнее осваивают свою экологическую нишу на (в) листьях зерновых культур при поражении корневой системы на 3–5 % и более возбудителями корневых гнилей. Это может быть объяснено активизацией гидролитических процессов, увеличением доли растворимых форм азотных и других соединений в растениях под влиянием возбудителей корневых гнилей (фузариозной, офиоболезной, обыкновенной).
Отношения типа синергизма при реализации экологических ниш вредными организмами возникает и в тех случаях, когда они занимают перекрывающиеся экологические ниши. Например, повреждения тканей корневой шейки внутристеблевыми вредителями открывает «ворота инфекции» для возбудителей гельминтоспориозной и фузариозной гнилей. При заражении корней картофеля нематодой (гетеродерой) возрастает зараженность их микозами, особенно F. oxysporum. В присутствии видов рода Verticillium на корнях растений перца синтез фузариевой кислоты у F. oxysporum возрастал в 25–40 раз. Грызуны стимулируют размножение двудольных растений (сорняков, бурьянов). Это обусловлено тем, что в результате поедания многолетних злаков, разнотравья, особенно вблизи нор (суслики, полевки), возникают лишенные растительности пятна, которые заселяются сорняками. Многие сорняки из таких сусликовин, кротовин переселялись на поля. На курганчиках малых сусликов поселяются белая полынь, злаки, достигающие больших размеров, чем на участках в культурных сообществах.
Многие вредители сначала питаются на сорняках, а затем на посевах культурных растений. Так, крестоцветные блошки вначале питаются на дикорастущих или сорных растениях семейства капустных, а затем переходят на всходы турнепса, брюквы, редьки, редиса, рассаду капусты. На диких растениях семейства капустных весной размножается капустная тля, которая затем переходит на капусту (табл. 25).
Таблица 25
Сорняки как источники питания для фитофагов
Вид сорняка |
Число фитофагов, питающихся на сорняках в Германии |
Пырей ползучий (Agropyron repens (L.) Pal. Beauv.) |
81 |
Мятлик однолетний (Poa annua L.) |
41 |
Горец птичий (Polygonum aviculare L.) |
40 |
Звездчатка средняя (Stellaria media (L.) Vill.) |
36 |
Горчица полевая (Sinapis arvensis L.) |
31 |
Горец почечуйный (Polygonum persicaria L.) |
28 |
Мать-и-мачеха обыкновенная (Tussilago farfara L.) |
25 |
Виды незабудки (Myosotis spp.) |
23 |
Редька полевая (Raphanus raphanistrum L.) |
23 |
Паслен черный (Solanum nigrum L.) |
17 |
Горошек волосистый (Vicia hirsuta (L.) Gray) |
16 |
Мак самосейка (Papaver rhoeas L.) |
15 |
Фиалка полевая (Viola arvensis Murr.) |
15 |
Торица полевая (Spergula arvensis L.) |
12 |
На бодяке полевом (Cirsium arvense (L.) Scop.) питаются 78 видов насекомых, из которых 6 строго монофаги, 5 только на нем и близкородственных видах, 26 живут только на растениях того же подсемейства и 44 вида более или менее специализированы, но живут только на сложноцветных. Число питающихся на пастушьей сумке (Capsella bursa-pastoris (L.) Medicus) фитофагов в средней Европе составляет 66 видов, из них 45 – более или менее олигофаги (виды, которые питаются только на нескольких видах) и 21 – полифаги (виды, питающиеся на многих видах).
Цветущие сорняки служат углеводным кормом для многих бабочек во время их дополнительного питания. Обилие цветов в период лета бабочек увеличивает их плодовитость и вредоносность.
Озимая совка откладывает яйца преимущественно на сорняки во второй половине лета, а развивающиеся гусеницы после этого заселяют озимые. В виноградниках в первой половине вегетации на щебенистых почвах сорняки не удаляют, так как они мульчируют почву, служат источником органики и местом скопления полезной энтомофауны. Это пример того, как сорные растения выступают в качестве полезного компонента биоценоза. Все подобные примеры свидетельствуют о том, что при реализации экологических ниш вредными организмами между ними в популяциях культурных растений возникают положительные взаимодействия, облегчающие этот процесс и дифференциацию экологических ниш.
При симбиозе обеспечивается тесное сотрудничество в пространстве и коадаптация взаимодействующих популяций. Пищевые отношения принимают характер комменсализма или мутуализма. При комменсализме одна популяция получает пользу, тогда как другой популяции это безразлично. Примером таких отношений могут служить эпифиты – микроорганизмы, обитающие на надземных органах растений за счет выделений их клеток и тканей и не причиняющие растениям вреда. Мутуализм характеризуется благоприятным взаимодействием популяций, полезным сожительством. Примером такого сожительства являются высшие растения и микроорганизмы ризосферы, цветковые растения и насекомые-опылители.
При совпадении и пересечении экологических ниш могут возникать антагонистические отношения антибиоза (несовместимости, антагонизма), ограничивающие или исключающие сосуществование видов. Такие отношения складываются между растениями и возбудителями болезней, а также среди бактерий и грибов, где взаимное вытеснение достигается с помощью выделения в среду особых антибиотических веществ.
В качестве антибиотических веществ организмы-консорбенты используются антибиотики, токсины и ферменты. Благодаря достижениям биотехнологии биопрепараты на их основе широко используются в растениеводстве, ветеринарии и медицине. При этом у возбудителей инфекционных болезней растений особая роль принадлежит токсинам, а у сапротрофов – антибиотикам. Фитопатогены выделяют токсины, ослабляющие растения, облегчая проникновение паразита в экологическую нишу, регулирующие проницаемость клеточных мембран растений, индуцируя отток из клеток и тканей питательных веществ, необходимых для жизнедеятельности возбудителей. В процессе эволюции возбудители болезней приобрели в процессе эволюции устойчивость к защитным реакциям хозяина (фенольные соединения, ферменты и др.), что делает их взаимоотношения на определенных этапах инфекционного процесса нейтральными и даже симбиотическими. Чем сильнее специализированы возбудители, тем выше совместимость в патосистеме растение-возбудитель.
Сапротрофы и хищники не имеют устойчивости к защитным механизмам хозяина, поэтому их существование внутри растений или животных более затруднительно.
Антагонисты подавляют конкурентов с помощью аллелохимических соединений: антибиотиков, ферментов, токсинов, которые выполняют роль репеллентов, отпугивающих другие виды, или убивающих конкурентов. В результате антибиоза происходит дифференциация экологических ниш организмов в сообществе в соответствии с принципами, сформулированными Г. Гаузе (см. раздел «Экологические ниши вредных организмов»).
Конкуренция является одной из форм антибиоза, ограничивающей возможность сосуществования видов в одной экологической нише экосистемы. Пищевая конкуренция часто сопровождается борьбой за пространство. Если два вида находятся на одном трофическом уровне, между ними весьма вероятна конкуренция за пищу. Со временем адаптивная эволюция одного или обоих видов может привести к тому, что каждый вид в пределах одного трофического уровня будет занимать отдельную экологическую нишу и конкуренция, таким образом, будет сведена к минимуму. В других случаях, когда конкурирующие виды занимают одинаковые или сильно перекрывающиеся ниши, может быть достигнуто равновесие, при котором ни один из видов не будет в массе размножаться.
В конкурентных взаимоотношениях культурных и сорных растений большая роль принадлежит аллелопатии (от греч. allelon – взаимно и pathos – страдание), то есть взаимовлияние консорбентов посредством выделения во внешнюю среду физиологически активных веществ: маразминов (вещества завядания), колинов (тормозящие развитие растений). Считают, что причина недостатка азота и калия на полях кукурузы, засоренных пыреем ползучим, связана с действием аллелопатических веществ пырея, снижающих поглощающую способность корней кукурузы и (или) переводящих питательные вещества в недоступную для нее форму. По этой же причине может снижаться способность пшеницы поглощать фосфор, даже если он имеется в почве в доступной форме и достаточном количестве.
Анализ экологических ниш позволяет понять причины неудач в использовании биометода, связанные с практическим применением микробных препаратов (на основе антагонистов и гиперпаразитов). |
Для подавления фитопатогена антагонистом необходимо, чтобы их экологические ниши совпадали хотя бы на некоторых фазах жизненного цикла. Например, совпадение экологических ниш Agrobacterium tumefaciens и A. radiobacter используется для подавления первого вида. Для повышения конкурентоспособности антагонистов в ризосфере растений необходимо освободить там экологические ниши для антагониста-интродуцента. Для этого используют штаммы, активно заселяющие ризосферу, способные войти в состав микоризы корней или эпифитов надземных органов растений.
По Г. Гаузе конкурентное исключение возможно только в случае полного или частичного совмещения экологических ниш патогена и его антагониста. Антагонисты (гиперпаразиты) могут воздействовать только на сапротрофную стадию (совмещение ниш или включение в трофическую цепь) и (или) на покоящиеся структуры патогена. Паразитическая стадия возбудителя находится под защитой растений и недоступна для антагонистов (гиперпаразитов). Под влиянием фунгицидов и других пестицидов происходит элиминация полезных видов ризосферы и филлопланы. При этом освободившиеся экологические ниши замещаются другими видами. В связи с этим важно знать сукцессию вредных организмов в экологической нише, обусловливающая последовательность ее заполнения. Это позволит предвидеть смену экологических ниш в сообществах.
Перекрывание экологических ниш у возбудителей листо-стеблевых инфекций Parastagonospora nodorum, S. tritici, Pyrenophora tritici-repentis может привести к расширению ниши у одного из видов при уничтожении пестицидами другого. Поэтому применение мер борьбы в отношении только одного вида может быть безрезультатно.
Тем самым конкуренция представляет собой любые взаимно отрицательные отношения, принимающие различный характер:
|



Взаимоотношения типа конкурентного антибиоза издавна привлекали внимание ученых. Так, Ч. Дарвин отмечал, что виды одного рода обычно сходны в своих потребностях, поэтому борьба между ними может быть более жесткой, чем между видами разных родов. Эта ситуация усугубляется при узкой специализации видов и узких размерах их экологических ниш. Первоначально конкуренция между видами описывалась уравнениями Лотки-Вольтерра, однако в современных условиях уточняется с учетом экологических ниш: если экологические ниши видов не пересекаются, то конкуренция между ними невозможна. При перекрытии экологических ниш один из видов элиминируется или же виды могут избегать конкуренции при существовании трофического благополучия в разных точках экологических ниш.
Специализация членов сообщества по питанию, использованию пространства, времени активности, сужает экологические ниши, а обратные процессы – их расширяют. На расширение экологических ниш действуют внутривидовая конкуренция, а на сужение – межвидовая типа «жертва – эксплуататор».
Отношения типа «эксплуататор – жертва» складываются тогда, когда увеличение (уменьшение) плотности популяции жертвы влечет за собой увеличение (уменьшение) скорости роста популяции эксплуататора, и наоборот, когда увеличение (уменьшение) плотности эксплуататора вызывает уменьшение (увеличение) скорости роста популяции жертвы. Наиболее важными примерами таких отношений являются система фитофаги – энтомофаги. Например, популяция гессенской мухи в типичные годы в значительной мере контролируется энтомофагами, в частности – паразитическими перепончатокрылыми (наездники). В суровые зимы из-за слабой морозоустойчивости энтомофаги гессенской мухи погибают практически полностью. Это позволяет вредителю восстановить численность и беспрепятственно размножаться вследствие отсутствия энтомофагов. По мере размножения гессенской мухи численность энтомофагов восстанавливается, также как и их регулирующая роль численности гессенской мухи.
При введении в экосистемы энтомофагов из 223 попыток полный успех контроля фитофагов был достигнут в 42 случаях, существенный – в 48, частичный – в 30 случаях. В остальных 103 случаях попытки интродукции энтомофагов были неудачными. Это обусловлено явлением «ускользания» жертвы от хищника, недостаточной адаптацией эксплуататора или, наоборот, полным уничтожением жертвы, что приводило к гибели эксплуататора. Относительно устойчивая ситуация складывалась в тех случаях, когда эксплуататор был достаточно эффективен против жертвы, быстро снижая ее численность до низкого устойчивого уровня, вблизи которого начинали действовать механизмы, препятствующие полному истреблению жертвы (наличие укрытий, иммунитет, ограничение контактов, изреженность популяции жертвы).
В качестве общего положения можно отметить, что вхождение в состав сообществ нового вида зависит в значительной мере от того, насколько его потенциальная экологическая ниша приближается к реализованной. В ненасыщенных сообществах, где ресурсы в цепях питания используются слабо или остаются свободными, возможно вхождение в сообщество энтомофагов с признаками r-стратегов. Успех проникновения вида в сложные насыщенные сообщества обычно зависит от его конкурентоспособности с аборигенами, занимающими близкую экологическую нишу. Если новый вид обладает достаточной конкурентоспособностью, происходит замещение экологических ниш одного вида другим, либо разделение одной ниши на две и усложнение сообщества при сужении экологических ниш. Во всех случаях в насыщенных сообществах новые популяции должны обладать признаками К-стратегов.
В природных условиях жизненные циклы эксплуататоров и жертв отрегулированы таким образом, что популяция жертвы никогда полностью не уничтожается, а лишь регулируется на определенном уровне, зависящем от емкости и сопротивления среды.
Под емкостью среды понимают предел, при достижении которого происходит ограничение численности вредных организмов в результате действия биотических и абиотических факторов, обеспечивающих сопротивление среды (рис. 12). |
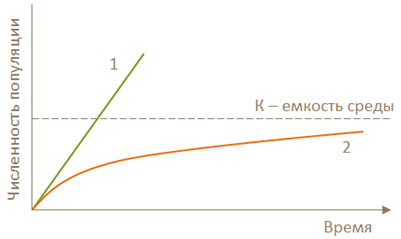
Рис. 12. Численность популяций при отсутствии сопротивления среды (1) и при ее наличии (2)
Сопротивление среды – это совокупность факторов, ограничивающих реализацию потенциальной способности популяций вредных организмов к росту численности в экосистемах. |
Для расчета сопротивления среды используют следующую формулу:
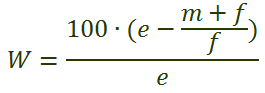
где: W – сопротивление среды, % гибели особей в популяции;
е – среднее число потомков одной самки;
m – число самцов;
f – число самок.
В начале роста численность популяций в условиях невысокой их плотности (численности) и хорошей обеспеченности ресурсами идет реализация биотического потенциала размножения с экспоненциальным ростом численности. Однако по мере увеличения плотности популяций, усиливается действие тормозящих рост внешних и внутренних факторов. В результате этого плотность поддерживается на некотором равновесном, свойственном данной экосистеме уровне, называемом «емкостью среды» по отношению к той или иной видовой популяции. Этот термин близок по смыслу к термину «емкость пастбища» в пастбищном хозяйстве, где емкостью пастбища называют максимальную плотность содержания животных, при которой еще сохраняется способность растительности к восстановлению, и таким образом, обеспечивается длительное поддержание высокого поголовья.
Данная схема описывает причины изменения численности популяций многих видов, но в некоторых случаях складывается более сложная картина. В частности, возможно, что сначала численность популяций в сообществах может колебаться беспорядочно. Но через несколько лет (около 70 поколений) вырабатывается динамическое равновесие на некотором среднем уровне численности (плотности).
В естественных экосистемах динамическое равновесие может сохраняться бесконечно долго, тогда как в агроэкосистемах оно нарушается хозяйственной деятельностью человека (антропогенными, или техногенными факторами). Для достижения равновесия в ненасыщенных наземных сообществах важна регулирующая роль трофических связей в системе триотрофа по схеме: растения – фитофаги – энтомофаги, а в насыщенных сообществах – освобождение в той же системе экологических ниш для антагонистов, хищников, обладающих признаками К-стратегов. Однако количественный уровень численности популяций при достигнутом равновесии в естественных и агроэкосистемах различен: в естественных он как правило ниже ПВ, а в современных агроэкосистемах – выше ПB и ЭПВ.
Для быстрого и радикального изменения ситуации в агроэкосистемах применяют пестициды, тем самым нарушая в большинстве случаев взаимоотношения и связи между консорбентами.
При экологическом направлении в защите растений важно задействовать долговременные механизмы саморегуляции агроэкосистем. Для этого необходимо учитывать следующие положения:
|



Для оптимизации фитосанитарного состояния агроэкосистем важно решить две задачи: |
|
|
сузить экологические ниши вредных организмов и расширить экологические ниши полезных и нейтральных: эпифитов, микоризообразователей, симбионтов, антагонистов, энтомофагов; |
|
повысить сопротивление (снизить емкость среды) для размножения, выживания, питания (тактик Р, В, Т) вредных организмов путем повышения активности антагонистов и энтомофагов, повышения супрессивности самой среды, в том числе устойчивости (генетической, индуцированной) и конкурентной способности культурных растений. |




