 Экологические основы интегрированной защиты растений: учебник
Экологические основы интегрированной защиты растений: учебник
 ЭКОЛОГИЧЕСКИЕ ОСНОВЫ ИНТЕГРИРОВАННОЙ ЗАЩИТЫ РАСТЕНИЙ
ЭКОЛОГИЧЕСКИЕ ОСНОВЫ ИНТЕГРИРОВАННОЙ ЗАЩИТЫ РАСТЕНИЙ

 ГЛАВА 1. ПОНЯТИЕ О СИСТЕМАХ ИНТЕГРИРОВАННОЙ ЗАЩИТЫ РАСТЕНИЙ
ГЛАВА 1. ПОНЯТИЕ О СИСТЕМАХ ИНТЕГРИРОВАННОЙ ЗАЩИТЫ РАСТЕНИЙ


3.2. Экологические ниши вредных организмов в агроэкосистемах
Реализация тактик размножения, выживания и трофических связей, а также жизненного цикла в целом происходит благодаря свойственным вредным организмам экологическим нишам.
Под экологической нишей в экологии понимают совокупность адаптаций, обеспечивающих особи, популяции или сообществу существование и воспроизведение потомства в экосистемах. |
На современном этапе развития эволюционного учения идея Ч. Дарвина о естественном отборе соединена с концепцией экологических ниш. Это служит основой для понимания стратегий жизненных циклов биологических видов в экосистемах.
Различают фундаментальные, или потенциальные экологические ниши и реализованные в тех или иных условиях окружающей среды. Фундаментальные ниши всегда шире реализованных.
Потенциальные возможности биологических видов к размножению при наличии доступных пищевых ресурсов и благоприятных условий чрезвычайно большие. Например, одуванчик за 10 лет способен заселить своими потомками весь Земной шар, если бы все его семена прорастали и давали новых потомков. Одна клетка бактерии, способной делиться каждые 20 мин, за 36 часов может дать потомство, которое сплошным слоем покроет всю нашу планету. Это не происходит из-за «сопротивления среды», или совокупного действия всех (физических, биологических и др.) факторов смертности в реальной среде, в результате чего организм не в состоянии реализовать свои потенциальные возможности размножения.
На границе компромисса между потенциальными потребностями организма и «сопротивлением» конкретной среды формируются в процессе эволюции экологические ниши, позволяющие занять популяциям вида определенное место и функционировать в сообществах экосистем.
Согласно закону конкурентного исключения Г. Гаузе, дифференциация (differentia – различие) экологических ниш подчиняется трем принципам:
|



Экологические ниши многомерны: они отличаются по многим параметрам – трофическим (пищевые ресурсы, место и тактика их использования), гидротермическим (зоны, годы, периоды вегетации), пространственным (ландшафты, географические ареалы) и т.д.
Выявленная Г. Хатчинсоном в 1957 г. многомерность экологических ниш считается новым этапом развития их теории.
Для вредных организмов важнейшим объектом среды первого порядка служат растения-хозяева, на (в) которых они размножаются, питаются и которые обеспечивают для них укрытия и убежища, защищая от неблагоприятных внешних условий окружающей среды, включая хищников и конкурентов.
Растения удовлетворяют основным требованиям, предъявляемым к экологическим нишам: они обеспечивают место существования вредных организмов, то есть их «дом» и возможность выполнения в экосистемах функций как биотических стрессов растений.
Именно биомасса (величина) органов популяции растений-хозяев определяют в значительной мере размер экологических трофических ниш вредных организмов в экосистемах. Считается, что ширина трофической ниши должна возрастать по мере уменьшения доступных ресурсов. В среде, где пищи мало, консумент не может игнорировать второстепенные пищевые ресурсы. Именно поэтому трофическая ниша расширяется, что приводит к широкой специализации вредных организмов. И, наоборот, в среде, богатой пищевыми ресурсами, время поиска пищевого объекта невелико. Вследствие этого игнорируются второстепенные пищевые ресурсы, происходит выборочное питание, рост специализации и сужение трофической ниши.
Таким образом, широкий уровень специализации в сочетании с поражением надземных вегетативных органов расширяет экологическую нишу, а узкая специализация в сочетании с поражением подземных или генеративных надземных органов сужает ее, но с повышением эффективности использования. Это обусловливает формирование различных экологических ниш вредных организмов в агроэкосистемах (рис. 7).
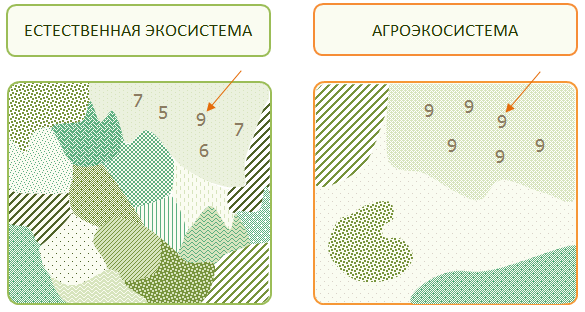
Рис. 7. Размер экологических ниш видовых популяций в естественных и агроэкосистемах
 характеристика (генетическая, возрастная, физиологическая) отдельных особей внутри популяции вида
характеристика (генетическая, возрастная, физиологическая) отдельных особей внутри популяции вида
В естественных экосистемах возрастает разнообразие особей в популяциях, в то время как в популяциях агроэкосистем они характеризуются относительной однородностью (генетической, возрастной, физиологической).
Кроме дифференциации экологических ниш в зависимости от состава фитоценозов в разных типах экосистем, они дифференцируются по другим параметрам, обусловленным специфическими эволюционными особенностями самих растений. Важнейшие из них заключаются в следующем:
 дифференциация тела растений на корневую систему, обитающую в почвенной среде и на надземные органы (лист, стебель, генеративные органы) с проводящими системами (ксилема, флоэма) структурно и функционально соединяющими подземные и надземные органы;
дифференциация тела растений на корневую систему, обитающую в почвенной среде и на надземные органы (лист, стебель, генеративные органы) с проводящими системами (ксилема, флоэма) структурно и функционально соединяющими подземные и надземные органы;
 защита зародыша семени от неблагоприятного влияния внешних условий благодаря обеспечению его питательными веществами, защитными покровами;
защита зародыша семени от неблагоприятного влияния внешних условий благодаря обеспечению его питательными веществами, защитными покровами;
 адаптация к распространению в пространстве спор и семян с помощью воздушных течений (анемохория), животных (зоохория), воды (гидрохория). Все возрастающую роль в распространении семян приобретает хозяйственная деятельность человека (антропохория).
адаптация к распространению в пространстве спор и семян с помощью воздушных течений (анемохория), животных (зоохория), воды (гидрохория). Все возрастающую роль в распространении семян приобретает хозяйственная деятельность человека (антропохория).
|
Расчленение тела растений на подземную и надземную части способствовало расхождению экологических ниш вредных организмов, приуроченности их к наземно-воздушной и (или) почвенной средам. |

Наземно-воздушная среда отличается по экологическим параметрам от почвенной. К наиболее существенным ее особенностям относятся:
 дефицит влаги. Режим увлажнения в наземно-воздушной среде разнообразен – от полного и постоянного насыщения воздуха водяными парами в областях экваториального и муссонно-тропического климата до практически полного их отсутствия в пустыне. В различных почвенно-климатических зонах Сибири, например, выпадает разное количество осадков: в степи – до 400 мм, в лесостепи – 400–500 мм, в предгорных и подтаежных областях – более 500 мм;
дефицит влаги. Режим увлажнения в наземно-воздушной среде разнообразен – от полного и постоянного насыщения воздуха водяными парами в областях экваториального и муссонно-тропического климата до практически полного их отсутствия в пустыне. В различных почвенно-климатических зонах Сибири, например, выпадает разное количество осадков: в степи – до 400 мм, в лесостепи – 400–500 мм, в предгорных и подтаежных областях – более 500 мм;
 колебания температуры. Суточные и годовые амплитуды составляют в Сибири, например, от –52°С до +45°С, а среднесуточные от –21°С (январь) до +20°С (июль). Для сравнения температура почвы на глубине 20 см зимой не снижается ниже –6°С, а на глубине более 3 м – всегда положительная;
колебания температуры. Суточные и годовые амплитуды составляют в Сибири, например, от –52°С до +45°С, а среднесуточные от –21°С (январь) до +20°С (июль). Для сравнения температура почвы на глубине 20 см зимой не снижается ниже –6°С, а на глубине более 3 м – всегда положительная;
 движения воздушных масс при более высокой интенсивности света и низкой плотности, которая создает незначительное сопротивление передвижению вредных организмов.
движения воздушных масс при более высокой интенсивности света и низкой плотности, которая создает незначительное сопротивление передвижению вредных организмов.
Почва служит твердой опорой для организмов и является источником разнообразных биофильных веществ (азотных, фосфорных, калийных, витаминов). По экологическим параметрам почва представляет собой промежуточную среду между наземной и водной. С наземной средой ее сближает наличие воздуха, относительно резкие изменения температуры и влажности верхних слоев. С водной средой почву сближает более стабильный в целом гидротермический режим, понижение содержания кислорода, присутствие в почвенных растворах солей и органических веществ. Несмотря на неоднородность экологических условий в различных типах почв, она характеризуется как достаточно стабильная среда, особенно для подвижных организмов. Почва, в отличие от воздушной среды, значительно в большей мере насыщена биотой, которая составляет 20–30 т/га в зонах с умеренным климатом. Корни растений составляют, примерно, 15 т/га, бактерии и актиномицеты – 4,5, грибы – 3, насекомые – 1, дождевые черви – 0,5 т/га.
Наземная среда и функционирующие в ней надземные органы растений служат основными экологическими нишами всех групп вредных организмов (табл. 20).
Таблица 20
Частота экологических ниш вредных организмов в различных средах, % [35]
Таксономическая группа |
Среда |
|
наземно-воздушная |
почвенная |
|
Вирусы |
97,8 |
2,2 |
Бактерии |
77,4 |
22,6 |
Грибы |
77,6 |
22,4 |
Насекомые: |
||
имаго |
100 |
34,1 |
яйцо |
65,9 |
42,0 |
личинка |
70,3 |
43,5 |
куколка и ложнокуколка |
28,4 |
73,4 |
Сорные растения |
||
многолетние |
85,0 |
100 |
однолетние (семена) |
100 |
100 |
На (в) клетках и тканях надземных органов растений возбудители вирусных, бактериальных и грибных инфекций размножаются, получая от них питательные вещества и энергию. Одновременно надземные органы служат для них укрытием от неблагоприятных условий внешней среды. При этом на разных этапах жизненного цикла в воздушной среде и в надземных органах растений споры грибных паразитов, например, адаптированы к разным условиям окружающей среды: при прорастании – к влажности, а в период заражения – к температуре.
Для реализации жизненной функции спор в наземной среде на фазе их прорастания особенно важна влажность, а в латентный (скрытый) период развития – оптимальная температура (для большинства видов 20–23 °С).
Насекомые-фитофаги на всех фазах их метаморфоза (кроме куколки) также адаптированы преимущественно к наземно-воздушной среде. Особенно высока связь с наземно-воздушной средой и надземными органами растений у имаго вследствие их функций – размножения и расселения в пространстве. Надземные органы служат экологической нишей для откладки яиц большинства насекомых-фитофагов (65,9 % из 200 широко распространенных видов), а также экологической нишей личинок, примерно, у 70,0 % насекомых-фитофагов. Личинки усиленно питаются надземными органами, растут и развиваются на (в) них, проходя периодически линьки со сбрасыванием кожной кутикулы.
В отличие от личинок, для куколок надземные органы являются менее благоприятной экологической нишей, вероятно, из-за больших перепадов влажности и температуры: только у 28,4 % насекомых куколки адаптированы к надземным органам растений (зонтичная моль, листовой люцерновый долгоносик фитономус, капустная белянка и др.).
Особенностью куколок, характерных только для насекомых с полным превращением, является их неспособность к питанию и пребывание в практически неподвижном состоянии. Они часто рассматриваются как стадия покоя, на протяжении которой происходят процессы внутренней перестройки личинки в имаго. Куколки очень часто защищены коконами, что в значительной мере повышает их выживаемость. Почва является более благоприятной средой для сохранения этой неподвижной стадии.
Дифференциация надземных органов растений способствовала дальнейшему расхождению экологических ниш вредных организмов. Одни используют в качестве экологической ниши листья (минирующие мухи, возбудитель столбчатой ржавчины смородины), другие – побеги однолетних культур, стволы деревьев (жуки-дровосеки, стеклянница), третьи – проводящую систему растений (возбудители трахеомикозов), четвертые – все надземные органы (возбудители антракнозов бобовых, саранчовые, луговой мотылек).
Кроме надземных органов, вредные организмы используют, хотя и реже, в качестве экологической ниши подземные органы: корневую систему, корне- и клубнеплоды. К ним относятся, например, возбудители корневых гнилей сельскохозяйственных культур – гельминтоспориозной, фузариозной, офиоболезной. В общей сложности из 350 относительно изученных возбудителей болезней почвенные фитопатогены составляют 20–25 %.
Бактерии и грибы используют подземные органы в качестве экологической ниши реже в 3–4 раза по сравнению с надземными органами, а вирусы – в десятки раз. При этом паразиты бактериальной и грибной природы приурочены преимущественно к заражению корне-клубнеплодов (возбудители обыкновенной, черной, порошистой парши картофеля).
Для имаго насекомых почвенная среда и растительные остатки на ней служат защитой от неблагоприятных условий, особенно в период летней и зимней диапауз. В большей мере с почвой связан жизненный цикл имаго у жуков. Например, серый свекловичный долгоносик забирается для зимовки в почву на глубину 20–50 см. Пройдя питание сначала на осоте, крапиве, чертополохе, вьюнке, а затем на всходах сахарной свеклы, подсолнечника или кукурузы, жуки после спаривания откладывают яйца в поверхностный слой почвы. Отродившиеся личинки и куколки развиваются также в почве. Широкие трофические связи и приуроченность всех фаз развития серого свекловичного долгоносика к почве привела к тому, что он утратил способность к полету и интенсивному освоению пространства. Однако подавляющее большинство жуков сохранили способность к полету (щелкуны, майский и июньский хрущи), хотя яйцекладка их производится в почву, в которой развиваются также личинки и куколки.
Фитофаги часто (в 42 % случаев из 200 широко распространенных вредителей сельскохозяйственных культур) откладывают яйца в почву около питающих растений, хотя реже, чем на надземные органы. Примерно с такой же частотой почву и подземные органы растений используют в качестве экологической ниши их личинки.
Жизнь личинок начинается после выхода из яйца. В этот период они, как правило, имеют относительно мягкие покровы, которые у открыто живущих личинок быстро окрашиваются и затвердевают, напоминая у щелкунов, например, отрезок проволоки, отсюда и название личинок – «проволочники».
В отличие от наземно-воздушной среды, почва служит основной топической экологической нишей для куколок насекомых. Многие насекомые не делают коконов в почве, окукливаясь в изготавливаемых округлых полостях, укрепляя их стенки экскрементами или другим способом, образуя тем самым земляные колыбельки (желтый тихиус-семяед, свекловичные блошки, люцерновый скосарь).
Сорные растения как свободноживущие виды значительно в большей степени адаптированы как к почвенной, так и наземной среде, что объясняет их широкое распространение и вредоносность, особенно при наличии широких экологических ниш в агроэкосистемах.
Адаптация к экологическим нишам в наземно-воздушной и почвенной средах предоставляет вредным организмам различные шансы для реализации основных тактик их жизнедеятельности.
Наземно-воздушная среда и функционирующие в ней надземные органы растений обеспечивают хорошие возможности для реализации тактики размножения вредных организмов, особенно половым способом. |
В условиях дефицита влаги в воздушной среде у грибов и частично сорных растений гаметофит сменился спорофитом, а у насекомых, нематод, грызунов наружно-внутреннее оплодотворение заменилось копуляцией. Самцы многих бабочек находят самок по запаху на расстоянии до 3 км, поэтому, возможно, они и летают против ветра при небольшой его скорости. Используются также зрительные, слуховые и биохимические сигналы для поиска партнеров в наземной среде. После внутреннего оплодотворения самки откладывают яйца, которые дают начало потомству.
Почвенная среда значительно в меньшей степени пригодна для размножения вредных организмов, особенно половым путем. У некоторых почвенных насекомых, например, двухвосток, подур оплодотворение не сопровождается контактом самцов и самок, то есть происходит без спаривания. Самцы рассеивают капельки спермы в местах обитания вида даже в отсутствие самок. Оплодотворение происходит при встрече самок со спермой. Следовательно, сперма находится сначала во внешней среде и лишь затем попадает без участия самца в половые пути самок, то есть во внутреннюю среду, что характерно для водных организмов.
Тактика выживания значительно более успешно реализуется в относительно стабильной почвенной среде. |
Формирующиеся специальные структуры у грибов (например, склероции, цисты), а также семена в толстых твердых оболочках у сорняков обеспечивают длительное (5–10 лет и более) выживание в почве, которая служит резервуаром вредных организмов в агроэкосистемах, особенно при повторных и бессменных посевах.
Трофические связи значительно шире у вредных организмов в наземно-воздушной среде. Наоборот, в почвенной среде трофические ниши вредных организмов довольно узкие.
Наземная среда создает благоприятные условия для расширения трофических ареалов особенно для подвижных вредных организмов в годы их размножения. Например, молодые малые суслики и песчанки передвигаются за сезон на 3–5 км, а длиннохвостый суслик может преодолеть расстояние в 180 км, особенно подвижны самцы.
Расселение и захват новых территорий у грызунов производится чаще созревающей молодью, которая очень активна. На периферийной части ареалов гибель особей значительно выше, чем в центре. Каждая популяция занимает «зону сходного благоприятствования» и их пространственная ниша в известной степени изолирована от соседних популяций. Однако у подвижных популяций границы ареалов периодически нарушаются.
Таким образом, у вредных организмов выработалась в процессе эволюции потребность в определенных размерах территории, которую называют жизненным пространством вредного организма. |
У вредных организмов, жизненный цикл которых связан с почвой, жизненное пространство в почвенном профиле измеряется, примерно 1–1,5 м в ширину и в глубину. У наземных вредных организмов оно гораздо шире, охватывая поля одной сельскохозяйственной культуры (шведская муха) или нескольких культур, например, гороха и многолетних бобовых трав (гороховая тля). Большую группу составляют вредные организмы, жизненное пространство которых охватывает агро- и естественные экосистемы. Например, свекловичные блошки в весенне-летний период питаются и размножаются на культурных и сорных растениях в агроэкосистемах (сахарная свекла, гречиха, ревень, щавель, щавелелистная гречишка), а осенью переселяются в места зимовок – в древесно-кустарниковые заросли, лесополосы, обочины дорог (естественные экосистемы). В связи с этим жизненное пространство свекловичных блошек включает пространство обоих типов экосистем.
Адаптация вредных организмов к определенным гидротермическим условиям в экологических нишах наземно-воздушной среды обуславливает смену их стаций в различных зонах (рис. 8).
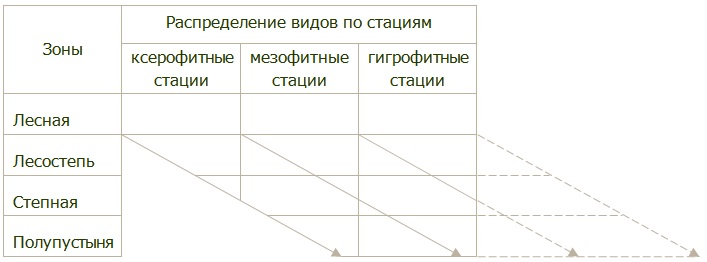
Рис. 8. Диаграмма зональной смены стаций (по Г.Я. Бей-Биенко)
Все стации по характеру растительного покрова и микроклимата разделяют на три экологические группы – ксерофитные (сухие теплые стации с разреженным травостоем), мезофитные (умеренно и среднеувлажненные) и гигрофитные (сильно увлажненные с густым травостоем). Смещение стаций происходит по зонам. Ксерофитная стация в лесостепи имеет такие же параметры как мезофитная в степной зоне. Это обусловливает сходные гидротермические условия для популяций в разных зонах: ксерофильные виды – такие как чернотелки, саранчовые, хлебные жуки – занимают более сухие, теплые стации, а гигрофильные (стеблевой мотылек, хлебные пилильщики) – более увлажненные, даже в таких засушливых зонах, как степь или полупустыня.
Подобные закономерности расхождения экологических ниш по гидротермическим и другим параметрам выявлены В.А. Чулкиной у возбудителей корневых гнилей зерновых культур – микромицета Bipolaris sorokiniana, и видов рода Fusarium – F. avenaceum, F. oxysporum, F. solani и др. Первый возбудитель адаптирован к экологической зоне относительно теплой и сухой, с повышенной солнечной радиацией и близкой к нейтральной реакцией среды, а вторые – к прохладной и увлажненной экологической нише с пониженной радиацией, достаточно высокой концентрацией углекислоты и низкой – кислорода, кислой реакцией среды. В связи с разными параметрами экологических ниш B. sorokiniana паразитирует чаще на органах, расположенных на границе почва-воздух (колеоптиль, эпикотиль, влагалища прикорневых листьев, основание стебля), в засушливые годы, а также в южных степных районах, а виды рода Fusarium – на первичных и вторичных корнях, в прохладные увлажненные годы и в более северных зонах – тайге и подтайге. Таким образом, несмотря на кажущееся совместное паразитирование возбудителей корневых гнилей зерновых культур прослеживается четкое расхождение и отсутствие перекрывания экологических ниш согласно закону конкурентного исключения Г. Гаузе.
Сложные жизненные циклы вредных организмов позволяют им занимать более чем одну трофическую экологическую нишу, что обусловлено у возбудителей болезней разными механизмами передачи возбудителей во времени и в пространстве. Нами предложено различать основные и дополнительные экологические ниши возбудителей почвенных, воздушно-капельных, семенных и трансмиссивных инфекций (табл. 21).
Таблица 21
Основные и дополнительные экологические ниши возбудителей инфекционных болезней в экосистемах
Экологические ниши |
Группы возбудителей инфекций |
|||
почвенных |
семенных |
воздушно-капельных |
трансимиссивных |
|
Корневая система и подземные органы |
+ |
(+) |
(+) |
|
Листья, стебли |
(+) |
+ |
(+) |
|
Генеративные органы |
(+) |
+ |
(+) |
(+) |
Проводящая система |
(+) |
(+) |
(+) |
+ |
+ основные экологические ниши
(+) дополнительные экологические ниши
Возбудители корневых, или почвенных, инфекций имеют в качестве основной экологической ниши подземные органы растений – корневую систему, корне-клубнеплоды.
Попадание почвенных фитопатогенов в основную экологическую нишу обеспечено их способностью к длительному выживанию в почве в виде покоящихся структур (склероции, цисты, ооспоры, хламидоспоры, зигоспоры). Под влиянием корневых выделений растений-хозяев покоящиеся структуры прорастают и заражают подземные органы. Однако, кроме передачи через почву, возбудители корневых инфекций могут передаваться из года в год дополнительно через семена и посадочный материал, а в течение вегетации – воздушными течениями и каплями дождя (возбудители обыкновенной корневой гнили зерновых культур, ризоктониоза картофеля). Дополнительным механизмам передачи возбудителей соответствуют дополнительные экологические ниши – листья, стебли, генеративные органы, а также проводящая система (фузариозное и вертициллезное увядание сельскохозяйственных культур).
Возбудители воздушно-капельных инфекций адаптированы к основной экологической нише в клетках и тканях листьев и стебля благодаря воздушно-капельному механизму передачи пропагативных структур (конидий, аскоспор, базидиоспор, пикноспор). Одновременно пропагативные структуры заражают дополнительно генеративные органы (возбудители септориозов, аскохитозов), а также проводящую систему (сосудистый бактериоз капусты), реализуя, таким образом, дополнительную экологическую нишу.
Основная экологическая ниша семенных инфекций обусловлена передачей возбудителей из года в год от материнских растений к дочерним через семена (возбудители головневых заболеваний зерновых культур).
Основной экологической нишей возбудителей трансмиссивных инфекции является проводящая система, в которую часто попадают вирусы с помощью насекомых-переносчиков, системно заражая растение, включая подземные органы (клубни).
Сложные жизненные циклы насекомых, особенно метаморфоз, позволяют разным фазам жизненного цикла одного вида решать разные задачи, обитая в различных экологических нишах. Если трофической нишей бабочек являются цветущие растение, где они питаются нектаром и пыльцой, то трофическая ниша гусениц – надземные органы, особенно листья растений, а куколки – почва. При этом бабочки выполняют функцию размножения и расселения популяций вида, гусеницы – быстрого накопления биомассы, роста и частично расселения, а куколки – превращения тела личинок во взрослое насекомое и выживания во времени (покоя). Бабочки имеют сосущий ротовой аппарат, а гусеницы – грызущий. Все эти адаптации повышают шансы популяций видов в более полном использовании ресурсов среды, а, следовательно, обеспечивают их размножение и выживание в экосистемах.
При адаптации к воздушной среде естественный отбор благоприятствует формированию у популяций видов признаков r-стратегов с более широкими экологическими нишами (надземные органы), а при адаптации к почвенной среде – признаков К-стратегов с более узкими экологическими нишами (корневая система, подземные органы). Популяции видов, адаптированные к обеим средам, обладают смешанными признаками и разными размерами экологических ниш.
Занимая различные экологические ниши, вредные организмы в меньшей мере конкурируют за ресурсы среды, формируя довольно стабильные сообщества в экосистемах. Лабильность экологических ниш вредных организмов более высока в наземно-воздушной среде, где возможны случаи их перекрывания и замещения.
|
В связи с этим применение мер борьбы с популяциями отдельных видов, занимающих сходные экологические ниши, малорезультативно. Возникает необходимость управления тактиками Р, В, Т всего комплекса вредных организмов – экологических эквивалентов. |

При этом у вредных организмов в наземно-воздушной среде важно нарушить адаптации тактики размножения, а в почвенной – выживания. Это будет способствовать снижению общего среднего уровня численности вредных организмов в обеих экологических средах агроэкосистем. Для обеих сред чрезвычайно важно сужение трофических и топических экологических ниш и замещение их симбиотрофными и нейтральными организмами.


